Amino acid metabolism follows a core logic of “amino-group transfer—deamination release/assimilation—carbon-skeleton recovery—resynthesis and allocation”, spanning protein turnover, energy metabolism, and nitrogen homeostasis. Key enzymes are frequently positioned at pathway entry points, irreversible steps, branch junctions, or intercompartmental coupling nodes. Their flux control is typically shaped by substrate accessibility, cofactor supply–demand balance, allosteric regulation, post-translational modifications, and expression/localization. A systematic organization of reaction essence, regulatory logic, compartmental constraints, and evidence-chain construction can map metabolite changes to testable biochemical steps, providing a reproducible research framework and experimental route for flux validation, mechanistic interpretation, and target intervention.
Keywords: amino acid metabolism; key enzymes; aminotransferases; glutamate dehydrogenase; branched-chain amino acid metabolism; threonine metabolism; D-amino acids; metabolic flux; stable isotope tracing; compartmentalization
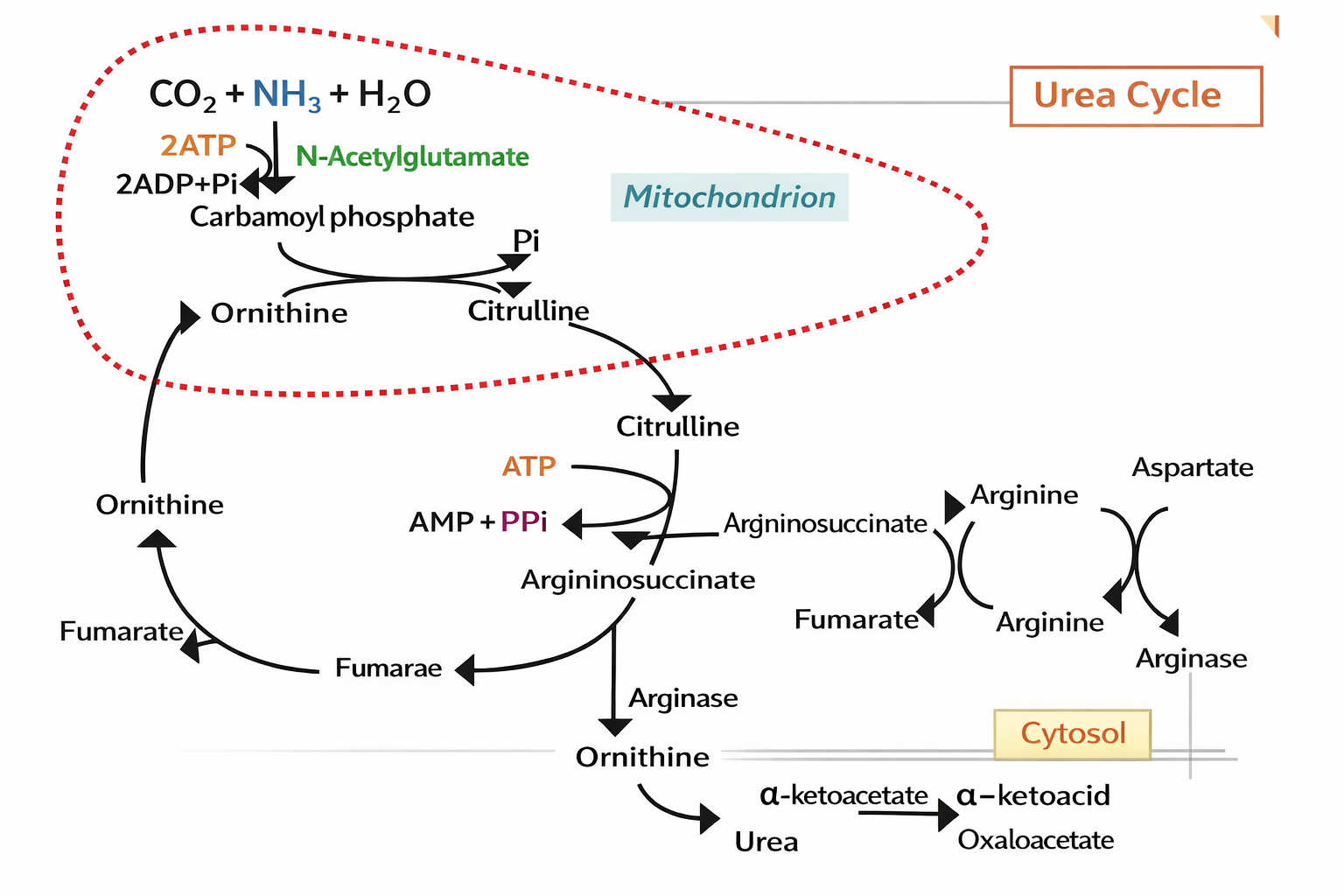
Figure 1 Schematic pathways associated with key enzymes in amino acid metabolism
I. Flux Control Points in Amino Acid Metabolism and the Logic for Defining Key Enzymes
1.1 Types of Research Questions and Evidence Hierarchy
(1) Existence and abundance level:
① Common evidence: targeted/untargeted metabolomics; amino acid analyzers; colorimetric/fluorometric quantification; key ratio metrics (e.g., Glu/Gln, BCAA/BCKA, urea/ammonia).
② Critical boundary: metabolite pool sizes reflect both production and consumption and should not be equated directly with flux changes. Extra caution is needed for fast-turnover, volatile, or readily recycled metabolites (e.g., NH4+, many α-keto acids).
(2) Flux and direction level:
① Common evidence: 13C/15N stable isotope tracing; isotopologue distribution analysis (M+0/M+1…); metabolic flux analysis models (MFA/INST-MFA); time-series comparisons of enrichment rates.
② Key value: upgrades “correlative changes” to “pathway-specific and directional evidence”, particularly suitable for reversible reactions and branch selection.
(3) Mechanism and regulation level:
① Common evidence: in-vitro enzyme activity/kinetics; verification of allosteric effects; detection of post-translational modifications; expression levels and subcellular localization; complex assembly states; key cofactors and energy-charge indicators (NAD(H), NADP(H), ATP/ADP/AMP) and mitochondrial functional readouts.
② Evaluation standard: should support a closed-loop explanation of “regulatory change—flux change—phenotypic change”, with causal validation via genetic/pharmacological perturbations when necessary.
1.2 Criteria for Defining Key Enzymes
(1) Entry-type key enzymes: determine whether a class of amino acids enters catabolic or anabolic networks, often co-limiting with transport and substrate supply.
(2) Branch-type key enzymes: determine allocation ratios of a shared substrate across different products or functional modules, often measurable as distinct isotope-flow patterns.
(3) Irreversible/rate-limiting key enzymes: set an upper constraint on pathway output and typically carry higher regulatory density and multi-layer regulation.
(4) Hub-coupling key enzymes: link central carbon metabolism (e.g., the TCA cycle) with nitrogen assimilation/clearance networks and shape carbon–nitrogen coupling; they often become structural constraints during metabolic reprogramming.
II. The Aminotransferase System: A Foundational Hub for Nitrogen Balance and Amino-Acid Interconversion
2.1 Reaction Essence and System-Level Significance
【Reaction type and cofactor dependence】
① Aminotransferases use pyridoxal phosphate (PLP) as a cofactor to catalyze amino-group transfer: “amino acid + α-keto acid ⇌ new amino acid + new α-keto acid”.
② Most transamination reactions are near-reversible; net direction is mainly determined by substrate/product concentrations, coupled reactions (subsequent deamination, anaplerosis, urea formation, etc.), and compartmentalization.
【Organization of nitrogen flow】
① Amino-acid catabolism often first concentrates amino groups onto hub amino acids such as glutamate, followed by deamination release or redistribution steps.
② Conversely, amino-acid biosynthesis and the synthesis of nitrogen-containing molecules use glutamate/glutamine as nitrogen donors to allocate nitrogen to diverse products.
【Key interpretation points for research】
① The transamination system behaves more like a nitrogen-exchange network than a one-way channel. Static abundance alone rarely resolves net direction; 15N tracing combined with isotopic pairing of keto acid–amino acid conjugate pairs is recommended.
2.2 ALT and AST: Coupled Reactions and Interpretation Boundaries
2.2.1 Alanine aminotransferase (ALT)
【Core reaction】
Amino-group transfer between alanine and α-ketoglutarate, generating pyruvate and glutamate.
【Metabolic coupling】
Links the pyruvate pool to the Glu/α-KG hub, directly influencing gluconeogenic substrate supply, the alanine cycle, and carbon–nitrogen coupling.
【Recommended evidence chain】
① Metabolites: alanine, pyruvate-related readouts, glutamate, α-KG, and glutamine.
② Tracing: 15N-alanine or 15N-glutamate for nitrogen flow; 13C-alanine for carbon-skeleton fate. Time-series enrichment helps distinguish enhanced exchange from increased net flux.
③ Enzymology: assess ALT activity and PLP availability to avoid misattributing cofactor limitation as flux regulation.
2.2.2 Aspartate aminotransferase (AST)
【Core reaction】
Amino-group transfer between aspartate and α-ketoglutarate, generating oxaloacetate and glutamate.
【Compartmentalization and energy coupling】
AST has cytosolic and mitochondrial isoforms and is tightly coupled to the malate–aspartate shuttle, influencing cytosolic NADH re-oxidation and coupling to mitochondrial oxidative metabolism.
【Recommended evidence chain】
① Metabolites: aspartate, glutamate, α-KG, and ratio information involving malate/aspartate/glutamate relevant to the shuttle.
② Tracing: combined 15N-aspartate with 13C-glucose or 13C-glutamate can help separate shuttle contributions from transamination contributions.
③ Localization: mitochondrial/cytosolic fractionation or isoform-specific detection to attribute flux changes to specific compartments.
III. Glutamate Dehydrogenase (GDH): A Key Gate for Deamination and Carbon-Skeleton Recovery
3.1 Reaction Modes and Network Position
【Core reaction and cofactor dependence】
① GDH catalyzes the reversible interconversion between glutamate and α-ketoglutarate with concomitant NH4+ release or utilization. The reaction may use NAD+ or NADP+, depending on species, isoforms, and cellular context.
② This node directly links the “nitrogen-concentrating pool (Glu)” with the “TCA-entry carbon skeleton (α-KG)”, making it a high-weight control point for carbon–nitrogen coupling.
【Testable physiological propositions】
① When carbon skeletons need to be recovered via the TCA cycle under changing nitrogen loads, GDH often channels flux in the Glu → α-KG direction and releases NH4+.
② Under appropriate metabolic background and cofactor supply–demand conditions, GDH may also contribute to NH4+ assimilation; net direction must be confirmed by tracing or flux evidence.
3.2 Coupling to Metabolic State and Regulatory Logic
【Allosteric regulation and linkage to energy status】
① GDH is commonly modulated allosterically by adenine nucleotides and related metabolites, coupling its activity to energy charge, carbon-skeleton demand, or nitrogen burden.
【Boundary of coupling to ammonia clearance modules】
① The consequences of GDH-mediated NH4+ release depend on the capacity of clearance pathways such as the urea cycle or glutamine synthesis. Interpreting GDH-related phenotypes therefore requires parallel assessment of NH4+ pools, glutamine synthesis, and urea-related readouts, to avoid misattributing “insufficient clearance” to “excess deamination”.
【Key interpretation points for experiments】
① Changes in Glu and α-KG abundance alone cannot establish GDH direction. Combine 15N tracing (nitrogen flow), 13C tracing (carbon-skeleton fate), enzyme activity assays, and cofactor/energy-charge readouts.
② For mitochondria-dominated GDH systems, include mitochondrial respiration, NADH re-oxidation capacity, and redox-state readouts to strengthen directional inference.
IV. Branched-Chain Amino Acid Metabolism: A Dual-Control Architecture of Entry Transamination and Downstream Oxidation
4.1 Entry Key Enzyme BCAT: Determines Whether BCAAs Enter the Catabolic Network
【Reaction positioning】
① Branched-chain aminotransferase (BCAT) catalyzes transamination between valine, leucine, and isoleucine and α-KG, producing the corresponding branched-chain α-keto acids (BCKAs) and generating glutamate.
② This step sets the entry threshold for net BCAA catabolism and is the key interface that merges the “BCAA pool” into the “keto-acid/oxidation module”.
【Research interpretation and application strategy】
① When BCAA abnormalities are observed, prioritize separating three mechanisms: entry limitation (BCAT or substrate accessibility), downstream limitation (oxidation-module bottlenecks), and changes in intake/transport.
② Entry limitation tends to present as elevated BCAAs without a marked rise in BCKAs; downstream limitation more often presents as BCKA accumulation with downstream product abnormalities.
③ Tracing recommendation: perform time-series tracing with 13C- or 15N-labeled BCAAs and quantify enrichment rates into BCKAs and further downstream products to localize the bottleneck directly.
4.2 Downstream Oxidation Module: A Common Control Point Setting the Upper Limit of Net Catabolic Flux
【System-level description and key-node logic】
① Further oxidation of BCKAs often constitutes a major limiting segment for net BCAA catabolism and may be regulated by phosphorylation and other mechanisms, producing a phenotype of “open entry but downstream congestion”.
② A more robust statement is that BCAA metabolism frequently exhibits joint control by entry and downstream modules; limitation at either segment can cause BCAA/BCKA accumulation. Flux evidence is required to identify the dominant bottleneck.
【Recommended evidence chain】
① Metabolites: parallel quantification and time-series trends of BCAAs, BCKAs, and characteristic downstream products to distinguish transient buildup from steady-state blockade.
② Enzymology and regulation: assess oxidation-module enzyme activities, key phosphorylation states, and mitochondrial oxidative capacity to explain changes in flux upper bounds.
③ Tracing: the transfer efficiency of enrichment from BCKAs to downstream products can serve as direct evidence for downstream bottlenecks.
4.3 Valine Dehydrogenase: Rigorous Positioning in Microbial Systems
【Functional summary】
① In some microbial contexts, oxidative deamination activity using valine as substrate can generate the corresponding α-keto acid and relate to nitrogen-source utilization and energy-acquisition strategies.
【Inference boundaries】
① Whether this node acts as a key rate-limiting point depends on species annotation, culture conditions, and flux allocation. It should be established by enzymology and tracing rather than extrapolated across systems.
V. The Threonine Metabolism Enzyme System: A Model Pathway for Branch Choice and Multi-Layer Regulation
5.1 Pathway Features and Why It Is Often Considered a “Regulatory Model”
【Multiple branches and strong coupling】
① Threonine has multiple branches in both biosynthetic and catabolic directions and couples to one-carbon metabolism, the serine/glycine network, and other amino-acid biosynthesis pathways.
② Consequently, its key enzymes often show regulation that is substrate-sensitive, feedback-rich, and branch-plastic, making the pathway suitable for studying general principles of branch selection and carbon–nitrogen coupling.
5.2 Key Enzyme Modules and Actionable Research Entry Points
【Transamination-related module】
① Controls redistribution of threonine-related amino groups and carbon skeletons across amino-acid pools, influencing nitrogen balance and steady states of the interconversion network.
【Cleavage/deamination-related module】
① Determines the capacity to divert threonine toward catabolism, often behaving as an “exit” flux-control feature and potentially changing coupling strength to one-carbon metabolism.
【Experimental strategies to determine branch choice】
① Static level: simultaneously monitor threonine, related keto acids/organic acids, and key one-carbon readouts (e.g., Ser/Gly ratio and methyl-donor-related indicators) to build branch fingerprints.
② Flux level: perform parallel tracing using 13C-threonine or 13C-glucose and compare the proportions and rates of label entering different downstream products to identify dominant branches.
③ Mechanistic level: validate in-vitro kinetics and feedback inhibition for candidate key enzymes, prioritizing feedback effects that hold within physiological concentration ranges. If post-translational modifications are involved, provide matched evidence across identical conditions for “modification—enzyme activity—flux”.
VI. Enzymes Related to D-Amino Acid Metabolism: Substrate-Spectrum Expansion and Metabolic Integration
6.1 D-Amino Acid Oxidoreductases/Dehydrogenases
【Reactions and significance】
① In some bacterial systems, these enzymes catalyze deamination of D-amino acids to generate α-keto acids, enabling carbon-skeleton recovery and coupling to nitrogen metabolism.
② This capability expands the usable substrate spectrum and may support ecological adaptation and nutrient competition.
【Recommended evidence chain】
① Substrate spectrum and specificity: in-vitro substrate screening and kinetic parameters.
② System contribution: combine growth phenotypes, tracing, and genetic perturbations to validate flux contribution under specific ecological/nutritional conditions.
6.2 D-Amino Acid Aminotransferases: Bringing D-Substrates into the General Nitrogen-Flow Network
【Functional positioning】
① Through transamination, D-amino-acid carbon skeletons and amino groups can be integrated into more general metabolic networks, often via “keto-acid intermediates—reamination” routes for redistribution.
② Use mechanism-neutral wording for “D/L interconversion”: a more robust statement is “metabolic integration and redistribution via intermediates”, avoiding claims of direct configurational inversion.
VII. Research Applications and Experimental Design: From a “Key-Enzyme Hypothesis” to an Evidence Closed Loop
7.1 Recommended Evidence Combinations
(1) Use metabolite profiling to localize abnormal nodes
① Quantify key amino acids, hub keto acids (e.g., α-KG), BCKAs, and NH4+ in parallel; report key ratios (Glu/Gln, BCAA/BCKA, etc.) to distinguish entry-accumulation patterns from downstream-blockade patterns.
(2) Use stable isotope tracing to determine flux and direction
① 15N tracing resolves nitrogen allocation and deamination contributions; 13C tracing resolves carbon-skeleton entry into the TCA cycle or branch products in terms of proportions and rate features.
② For reversible reactions and branch networks, time-series sampling is recommended over single time points; use enrichment rates rather than enrichment levels alone as key evidence for directional inference.
③ When necessary, adopt dual-label strategies (13C + 15N) to lock carbon–nitrogen coupling and reduce ambiguity from a single isotope.
(3) Validate enzyme activity and regulation to explain mechanism
① Perform in-vitro activity and kinetic measurements for GDH, ALT/AST, BCAT, and downstream oxidation modules, and verify allosteric effects and post-translational modification states when required.
② Assess cofactors and energy charge (NAD(H), NADP(H), ATP/ADP/AMP) and mitochondrial functional readouts in parallel to explain why flux can change despite similar expression levels.
(4) Establish causality by perturbation
① Genetic perturbations: knockdown/knockout/overexpression or isoform-specific interventions to validate compartmental contributions.
② Pharmacological perturbations: inhibitors or activators to test flux reversibility; combine dose–response and rescue experiments to strengthen causality.
③ Phenotypic readouts: relate flux changes to cell proliferation, oxidative stress, ammonia toxicity readouts, or energy-status indicators.
7.2 Key Points for Isotope-Tracing Data Interpretation
(1) Natural-abundance correction and quality control
① Apply natural-abundance correction to isotopologue distributions and report internal standards, drift correction, and reproducibility metrics to avoid mistaking technical noise for flux differences.
(2) Distinguish exchange from net flux
① Reversible reactions often show rapid label exchange, causing enrichment increases without necessarily increasing net flux. Time-series enrichment rates and label transfer into downstream products better reflect net flux.
(3) Parallel branches and pathway disentanglement
① When a metabolite can be produced through multiple pathways, a single tracer can be ambiguous. Use dual-substrate tracing, complementary labeling positions, or parallel quantification of key branch products to split contributions.
(4) Compartmentalization-driven “false averages”
① Whole-cell measurements can average mitochondrial and cytosolic pools and mask true bottlenecks. When conclusions depend on compartment attribution, add fractionation, localization, or compartment-specific isotope evidence.
7.3 Engineering and Tool-Oriented Applications
(1) Metabolic engineering
① Enhancing nitrogen concentration/allocation nodes can increase synthesis efficiency of nitrogen-containing products, but side reactions, cofactor burden, and growth costs must be evaluated to avoid converting flux gains into toxicity or imbalance.
② For dual-control architectures (e.g., BCAA metabolism), engineering should favor coordinated modifications; single-node enhancement may simply cause intermediate accumulation.
(2) Enzymology tools and screening platforms
① Key enzymes can serve as flux indicators, inhibitor-screening targets, or metabolic-state probe candidates. This requires clarity on reaction specificity, compartment context, and system compatibility, with tracing-based validation of predictable flux effects.
VIII. Safety Notes and Data-Interpretation Considerations
8.1 Safety Notes
(1) Biosafety
① When working with cells, tissues, or blood-derived samples, follow the relevant biosafety level requirements, including PPE, aerosol control, and regulated waste disposal.
(2) Chemical safety
① Metabolomics workflows often involve organic solvents, derivatization reagents, and corrosive solutions. Operate under proper ventilation and prepare emergency procedures for spills, burns, and exposure.
8.2 Interpretation Considerations
(1) Pool size is not flux
① Metabolite increases can result from increased production or decreased consumption; establish directionality with tracing and/or enzyme-activity evidence.
(2) Cross-species extrapolation must be constrained
① A key node in microbes is not necessarily rate-limiting in mammals; rely on species annotation, expression/localization, and measured flux evidence.
(3) Reversible reactions and branch nodes require time dimension
① Single-time-point snapshots are more prone to misinterpretation for reversible/branch reactions; time-series tracing and kinetic sampling improve resolution.
IX. Quick Reference Table of Key Enzymes in Amino Acid Metabolism
Enzyme | CAS | Key Reaction | Pathway Node | Functional Notes | Common Applications |
Alanine aminotransferase ALT (EC 2.6.1.2) | 9000-86-6 | Alanine + α-ketoglutarate → Pyruvate + Glutamate | Alanine metabolism; nitrogen transfer hub | Channels amino-group nitrogen into the glutamate pool, linking amino acid catabolism with central carbon metabolism | Transamination assay systems; metabolic flux studies |
Aspartate aminotransferase AST/GOT (EC 2.6.1.1) | 9000-97-9 | Aspartate + α-ketoglutarate → Oxaloacetate + Glutamate | Aspartate metabolism; TCA interface | Couples nitrogen flow to TCA intermediates, supporting anaplerosis and nitrogen redistribution | Metabolic flux analysis; enzyme activity assays |
Glutamate dehydrogenase GDH (EC 1.4.1.3) | 9029-12-3 | Glutamate ⇄ α-ketoglutarate + NH₃ | Glutamate–αKG hub | A key valve for carbon–nitrogen coupling, controlling ammonia release and αKG replenishment | Ammonia production/consumption assays; αKG-coupled systems |
Glutamine synthetase GS (EC 6.3.1.2) | 9023-70-5 | Glutamate + NH₃ + ATP → Glutamine | Core nitrogen assimilation | Converts free ammonia into glutamine, supporting multiple biosynthetic pathways | Nitrogen metabolism models; glutamine supply/assimilation studies |
Glutaminase GLS (EC 3.5.1.2) | Glutamine + H₂O → Glutamate + NH₃ | Entry to glutamine catabolism | Generates glutamate and releases ammonia, driving glutamine catabolic flux | Cancer metabolism; ammonia-release evaluation | |
Asparaginase ASNase (EC 3.5.1.1) | Asparagine + H₂O → Aspartate + NH₃ | Asparagine catabolism | Lowers asparagine while producing aspartate, altering nitrogen availability | Asparagine-dependence models; ammonia-release assays | |
Arginase ARG (EC 3.5.3.1) | 9000-96-8 | Arginine + H₂O → Ornithine + Urea | Terminal step of the urea cycle | Produces urea and ornithine, linking nitrogen disposal to polyamine precursor supply | Urea production assays; arginine-depletion models |
Ornithine decarboxylase ODC (EC 4.1.1.17) | 9024-60-6 | Ornithine → Putrescine + CO₂ | Rate-limiting step in polyamine biosynthesis | Diverts ornithine toward polyamines, influencing growth and nitrogen partitioning | Polyamine metabolism; proliferation-associated studies |
Glutamate decarboxylase GAD (EC 4.1.1.15) | 9024-58-2 | Glutamate → GABA + CO₂ | GABA shunt | Redirects glutamate into the GABA pathway, reshaping carbon flux distribution via the shunt | GABA metabolism; neuroscience-related studies |
Serine hydroxymethyltransferase SHMT (EC 2.1.2.1) | 9029-83-8 | Serine + THF ⇄ Glycine + 5,10-methylene-THF | One-carbon metabolism interface | Supplies one-carbon units, affecting nucleotide biosynthesis and methylation capacity | One-carbon metabolism; isotope tracing |
Cystathionine β-synthase CBS (EC 4.2.1.22) | Homocysteine + Serine → Cystathionine | Entry to transsulfuration | Routes homocysteine into cysteine biosynthesis, impacting glutathione precursor supply | Sulfur amino acid metabolism; redox studies | |
Cystathionine γ-lyase CGL/CTH (EC 4.4.1.1) | Cystathionine → Cysteine + α-ketobutyrate + NH₃ | Downstream transsulfuration | Produces cysteine and α-ketobutyrate, connecting sulfur metabolism with energy metabolism | Cysteine metabolism; stress/redox models | |
S-adenosylmethionine synthetase MAT (EC 2.5.1.6) | Methionine + ATP → SAM | Methionine cycle | Determines SAM availability and thus global methyl-donor capacity | Methylation capacity; one-carbon metabolism | |
Methionine synthase MS (EC 2.1.1.13) | 9033-23-2 | Homocysteine → Methionine | Methionine regeneration | Maintains methionine and SAM supply; influences homocysteine levels | Methionine cycle; homocysteine-related studies |
Histidine ammonia-lyase HAL (EC 4.3.1.3) | 9013-75-6 | Histidine → Urocanate + NH₃ | Entry to histidine catabolism | Initiates histidine degradation and releases ammonia, affecting nitrogen balance | Histidine catabolism; ammonia-release evaluation |
Tyrosine aminotransferase TAT (EC 2.6.1.5) | 9014-55-5 | Tyrosine + αKG → p-hydroxyphenylpyruvate + Glutamate | Entry to aromatic amino acid catabolism | Channels tyrosine into degradation while transferring nitrogen to glutamate | Aromatic amino acid metabolism; hepatic metabolism models |
Tryptophan hydroxylase TPH (EC 1.14.16.4) | 9037-21-2 | Tryptophan → 5-hydroxytryptophan | Serotonin-axis branching | Controls flux partitioning of tryptophan toward the serotonin pathway | Serotonin-related studies; pathway flux analysis |
Phenylalanine ammonia-lyase PAL (EC 4.3.1.24) | Phenylalanine → Cinnamate + NH₃ | Deamination module | A representative deaminase used for catabolism studies and biocatalytic entry reactions | Biocatalysis; deamination reaction setup | |
L-amino acid oxidase LAAO (EC 1.4.3.2) | L-amino acid + O₂ → α-keto acid + NH₃ + H₂O₂ | General oxidative degradation module | Produces α-keto acids and H₂O₂, enabling convenient coupled readouts | Oxidative degradation; H₂O₂-coupled detection | |
D-amino acid oxidase DAAO (EC 1.4.3.3) | D-amino acid + O₂ → α-keto acid + NH₃ + H₂O₂ | D-amino acid metabolism | Specific for D-enantiomers; widely used for discrimination and quantification | D-amino acid analysis; biosensing/coupled assays | |
Tryptophanase TnaA (EC 4.1.99.1) | 9024-00-4 | Tryptophan → Indole + Pyruvate + NH₃ | Microbial tryptophan catabolism | Generates indole and pyruvate, linking aromatic amino acid breakdown to central carbon metabolism | Indole production; microbial metabolism studies |
Aspartate ammonia-lyase Aspartase (EC 4.3.1.1) | 9027-30-9 | Aspartate → Fumarate + NH₃ | Aspartate catabolism; anaplerosis | Directly supplies a TCA intermediate while releasing ammonia | Anaplerosis/flux studies; fumarate production |
Alanine–glyoxylate aminotransferase AGXT (EC 2.6.1.44) | 9015-67-2 | Alanine + Glyoxylate ⇄ Pyruvate + Glycine | Glycine–glyoxylate node | Connects glycine and central carbon metabolism; impacts glyoxylate clearance | Glycine/glyoxylate metabolism; transamination network reconstruction |
Serine dehydratase SDH (EC 4.3.1.17) | 9014-27-1 | Serine → Pyruvate + NH₃ | Entry to serine catabolism | Routes serine carbon skeleton directly into pyruvate, coupling amino acid catabolism to energy metabolism | Serine catabolic flux; ammonia and pyruvate readouts |
Lysine decarboxylase LDC (EC 4.1.1.18) | 9024-76-4 | Lysine → Cadaverine + CO₂ | Amine-product branching | Diverts lysine toward amine products, altering nitrogen allocation and amine balance | Amine product synthesis; microbial amine metabolism |
D-cysteine desulfhydrase (EC 4.4.1.15) | 84012-74-8 | D-cysteine → reduced-sulfur products + NH₃ | Sulfur catabolic branch | Participates in sulfur amino acid degradation and reduced-sulfur (e.g., H₂S-related) metabolism | Sulfur metabolism; reduced-sulfur/H₂S-related studies |
Key enzymes in amino acid metabolism can be interpreted systematically using a framework of “aminotransferase-organized nitrogen flow—nitrogen release/assimilation at nodes such as GDH—carbon skeleton entry into central metabolism—branch control shaping product spectra”. The aminotransferase system provides a general mechanism for nitrogen redistribution; ALT/AST and compartment coupling reveal constraints of carbon–nitrogen exchange and shuttle operation; GDH functions as a carbon–nitrogen gate whose net consequences are jointly determined with ammonia-clearance capacity; BCAA metabolism exemplifies a dual-control architecture of entry and downstream modules; threonine metabolism illustrates branch regulation and feedback features; and D-amino-acid-related enzymes reflect substrate-spectrum expansion and metabolic integration mechanisms. With flux-evidence priority, closed-loop mechanistic validation, and clear statement boundaries, key enzymes can be advanced from conceptual nodes to actionable and reproducible research and application pathways.
For more related articles, please see below:
[1] Analysis of amino acid metabolites
Aladdin: https://www.aladdinsci.com/