Summary
Yeast two-hybrid (yeast two-hybrid) is an experimental system for studying the effects of interactions between trans-acting factors on the transcriptional regulation of eukaryotic genes based on yeast genetic analysis.
Its most valuable application is the screening of cDNA libraries composed of AD-Y with BD-X for novel protein interactions and the analysis of new genes for study.
This technique can be used not only to identify new protein interactions, confirm suspected interactions, and determine the structural domains of interactions, but also to directly obtain genes encoding interacting proteins.
Principle
The basic principle of yeast two-hybrid technology is that many transcription factors contain two independent functional domains, i.e., the DNA binding domain (BD) and the transcription activation domain (AD). Transcription factors bind to specific sequences on DNA through BD and AD, respectively, to initiate transcription of the corresponding genes.
The yeast two-hybrid system first constructs two trans-acting factors by fusing protein X with a reporter gene transcription factor-specific BD (e.g., Gal4-BD, LexA-BD) to become the bait (bait), and protein Y with a specific AD (Gal4-AD, B42-AD) to become the prey (prey).
When the genes encoding the two structural domains are simultaneously expressed in the yeast nucleus, if there is a non-covalent interaction between proteins X and Y, the upstream ac-tivation sequence (UAS) of the two structural domains of AD and BD will be close to each other, which in turn will activate the transcription process and enable the reporter genes (e.g., HIS3, LEU, and lacZ, etc.) to be expression (Fig.)
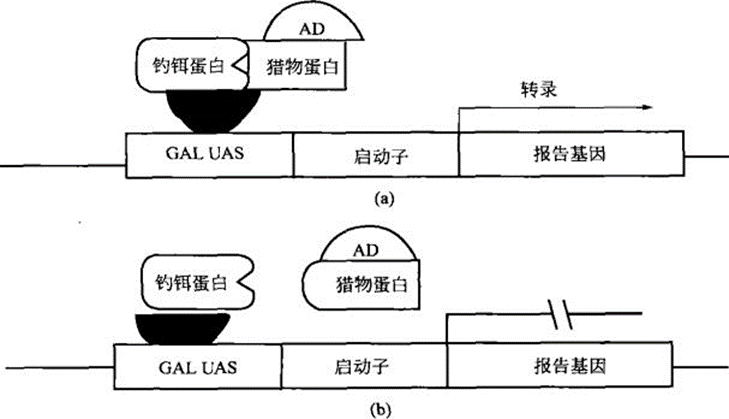
The connection (interaction) between BD and AD can effectively activate transcription. Two structural domains connected by a non-covalent bond can also connect BD and AD through protein-protein interactions, thereby initiating transcription; conversely, the presence of interactions between structural domain-connected proteins can also be evaluated by reporter gene expression.
Appliance
The most valuable application of the yeast two-hybrid system is the screening of cDNA libraries consisting of AD-Y with BD-X to obtain new protein interactions and to analyze and study new genes. This technique can be used not only to identify new protein interactions, confirm suspected interactions, and determine the structural domains of interactions, but also to directly obtain genes encoding interacting proteins.
Operation method
Yeast two-hybrid assay
Principle
The basic principle of yeast two-hybridization is that many transcription factors contain two independent functional domains, i.e., the DNA binding domain (BD) and the transcription activation domain (AD). Transcription factors bind to specific sequences on DNA through BD and AD, respectively, to initiate transcription of the corresponding genes. The yeast two-hybrid system first constructs two trans-acting factors by fusing protein X with a reporter gene transcription factor-specific BD (e.g., Gal4-BD, LexA-BD) to become bait, and protein Y with a specific AD (Gal4-AD, B42-AD) to become prey. When the genes encoding the two structural domains are simultaneously expressed in the nucleus of yeast cells, if there is a non-covalent interaction between proteins X and Y, the upstream ac-tivation sequence (UAS) of the AD and BD structural domains will be close to each other, which will activate the transcriptional process, and the reporter genes (e.g., HIS3, LEU, and lacZ, etc.) are expressed (as shown in the figure). The linkage (interaction) between BD and AD can effectively activate transcription. Two structural domains connected by a non-covalent bond can also connect BD and AD through protein-protein interactions to initiate transcription; conversely, the presence of interactions between domain-connected proteins can be evaluated by reporter gene expression.
Materials and Instruments
Equipment:
① Electrothermal constant temperature incubator
② Ultra-clean bench
③ tabletop high-speed centrifuge, high-speed freezing centrifuge
④ thermostatic water bath, thermostatic water bath shaker
⑤ autoclave, sterilization with filters
⑥pH meter
(vii) Electroconversion apparatus, electroconversion cups
⑧ vortex mixer
⑨ various specifications of conical flasks, 50 ml centrifuge tubes, 15 ml centrifuge tubes,
1.5 ml Eppendorf tubes, 100 cm Petri dishes, cellulose acetate membrane or filter paper.
Reagents:
①Materials: yeast AH109 or Y187, E.coli TG1, cDNA library
Tryptone, yeast extract, agar, glucose, various amino acids, yeast nitrogen base.
③ NaCl, PEG4000
③ NaCl, PEG4000 ④ Galactopyranoside derivative (5-bromo-4-chloro-3-indolyl-α-D-galactopyranoside, X-a-gal) or X-β-gal
⑤ 1 mol/L 3-amino-triazole (3-amino-1,2,4-triazole, 3-AT)
⑥ Fish sperm DNA
⑦ DMSO, glycerol
⑧ Lithium acetate
⑨ SDS, Lyticase
⑩ Phenol-chloroform (1:1), anhydrous ethanol, etc.
Move
The basic process of yeast two-hybridization can be divided into the following steps:
1. Preparation of yeast receptor cells and transformation of vectors (small-scale transformation)
(1) Pick the fresh yeast AH109 monoclonal clone from the YPD plate, which has been grown for 1~2 weeks and has a diameter of 2~3 mm, and inoculate it into 1 ml of sterilized deionized water, and shake it vigorously to disperse the yeast mass.
(2) Transfer the above yeast cells to 30 ml of YPD culture medium.
(3) Cultivate the yeast cells at 30 ℃ and 250 r/min overnight until stabilization (OD600>1.5).
(4) Transfer appropriate amount of the above cultures into 150 ml of YPD culture medium until the OD6oo reaches 0.2~0.3, and incubate at 30 ℃ and 250 r/min until the OD600 reaches 0.5±0.1.
(5) Transfer to a 50 ml centrifuge tube and centrifuge at 4000 g for 5 min at room temperature, discard the supernatant. Resuspend the cells in 25-50 ml of sterilized TE or distilled water with vigorous shaking. Repeat centrifugation and discard the supernatant.
(6) Resuspend the precipitate in 0.75 ml 1xTE/LiAc, this is the yeast receptor cell, and set aside in an ice bath.
(7) Add the following ingredients into a 1.5 ml Eppendorf tube and shake well.
(8) Add 0.1 ml of yeast receptor cells and shake well.
(9) Add 0.6 ml of sterilized PEG/LiAc solution and mix with high speed shaking.
(10) Cultivate at 30 ℃ and 200 r/min for 30 min.
(11) Add 70 μl of DMSO, gently invert up and down and mix well (can not be shaken vigorously). 42 ℃ heat shock for 15 min, and immediately cooled in an ice bath for 2 minutes.
(12) Centrifuge at 4000 g for 5 min and discard the supernatant as much as possible. Resuspend the cells in 0.5 ml of 1xTE (pH 7.5).
2. Spreading and screening of the transformation mixture
(1) Spread the above transformation mixture on a plate of appropriate medium (100 mm diameter Petri dish) as shown in Figure 12-5. It is difficult to predict the most suitable method for combining a new bait with a library.
Therefore, 1/3 of the transformants were spread on different selection media: low rigor (SD/-Leu/-Trp), medium rigor (SD/-His/-Leu/-Trp) and high rigor (SD/-Ade/-His/-Leu/-Trp/X-a-gal) media.
(2) Invert the culture at 30 ℃ until the clones grow out, and the clones that show blue color on X-a-gal medium are positive. The β-galactosidase filter blotting method can also be performed without adding X-a-gal according to step (3).
(3) Determination of positive clones by β-galactosidase membrane blotting assay
Prepare Z buffer/X-β-gal solution.
② with tweezers, a dry sterilized nitrocellulose filter membrane (or filter paper) placed on the above colony plate, gently press the membrane with tweezers so that the clone adheres to the filter membrane.
③ When the filter membrane is evenly wetted, carefully remove the membrane from the medium and place it in liquid nitrogen.
④After being completely immersed in liquid nitrogen for about 15 s, remove the membrane and place the clone side up into a clean petri dish (or lid).
⑤ Add 1-2 ml of Z buffer/X-β-gal solution until the membrane is completely wetted.
⑥ Incubate at 30 ℃. Check the appearance of blue clones within 8h.
3. Identification and confirmation of positive clones
(1) Inoculate the galactosidase-positive clones onto SD/Leu/Trp/X-α-Gal discs; or inoculate them onto SD/Leu/Trp discs, and then carry out the β-galactosidase blotting method for the identification of the grown clones. Meanwhile, the positive clones were stored at -70 ℃.
(2) Invert the culture at 30 ℃ for 4~6 d. Observe the color change of the clones or use the β-galactosidase membrane blotting assay to show the color change.
(3) Repeat the above steps (1)-(2) two to three times.
4. Yeast plasmid isolation
(1) Amplify a certain amount of yeast on a positive clone yeast disk, scrape off about 30 μl, add 50 μl of TE (pH 7.0), and vortex and shake to suspend the cells.
(2) Add 10 μl of Lyticase lysate (5 U/μl) and mix by vortexing or repeated pipetting.
(3) Incubate at 37 ℃ for 1 h at 250 r/min with shaking.
(4) Add 10 μl of 20% SDS and vortex for 1 min.
(5) Freeze (-20 ℃)/thaw once and vortex to fully lyse the cells.
(6) Make up 200 μl with TE.
(7) Add 200 μl of phenol-chloroform (1:1) and vortex at high speed for 5 min.
(8) Centrifuge at 10,000 g for 10 min and transfer the aqueous phase to a new Eppendorf tube.
(9) Add 1/10 volume of 5 mol/L NH4Ac solution and twice the volume of anhydrous ethanol.
(10) Freeze at -70 ℃ for 1 h. (11) Centrifuge at 10,000 g for 10 h.
(11) Centrifuge at 10,000 g for 10 min and discard the supernatant.
(12) Dry the DNA precipitate and dissolve it in 20 μl of sterilized water.
5. Electrotransformation of yeast plasmid
(1) Add 2-5 μl of yeast plasmid into 40 μl of receptive yeast cells, mix well and ice bath.
(2) Transfer the mixture into an ice pre-cooled electrochemical cup, set appropriate parameters (e.g. 1.8 kV, 25 μF, 200 Ω), and electrocute.
(3) Quickly transfer the cytosol into 1 ml of SOC culture medium and mix gently. incubate at 37 ℃ and 250 r/min for 1h.
(4) Centrifuge at 4000 g for 5 min, discard the supernatant, keep about 100 μl of culture medium, gently resuspend and spread on LB/Amp Petri dish plate, and incubate inverted at 37 ℃ overnight.
The bacterial plasmid was extracted according to the conventional method. The extracted plasmids were subjected to small-scale transformation experiments to confirm the interaction between the positive clones and known genes.
6. Classification of positive clones
Plasmids are digested with restriction endonucleases inserted at both ends of the cDNA fragments, and the size of the inserted fragments is identified by agarose gel electrophoresis and categorized accordingly.
7. Cotransformation of representative clones with lures
A representative clone from each category was co-transformed with the bait to confirm whether it actually interacts with the bait in yeast (the method is the same as that of the small-scale transformation experiment).
8. Sequence the plasmids of the confirmed positive clones.
9. Blast the sequences for comparison. In case of unknown cDNA sequences, additional bioinformatics analysis can be performed (www.ncbi.nlm.nih.gov/blast).
10. The obtained interacting protein pairs need to be confirmed by other methods, such as immunoprecipitation, GST-pull down, etc.
Caveat
(1) Small-scale transformation experiments are commonly used to: confirm DNA-BD/trolls that do not self-activate reporter genes, confirm the toxicity of DNA-BD/trolls to the host, control experiments, and sequential transformation of DNA-BD/trolls.Sequential transformation means that the DNA-BD/jig plasmid is first introduced into yeast by small-scale transformation, and then the AD-fused library plasmid is transferred into selected yeast clones.(2) Generally extracted yeast plasmids are suitable for electrotransformation since it is difficult to obtain transformed bacterial clones by chemical transformation method due to low DNA content and mixed impurities.For electroconversion, different settings should be tried to find the right parameters.(3) In β-galactosidase membrane blotting, the clones should be 1-3 mm in diameter. If there are only a few clones on each plate, the clones can be grouped together on a master plate.In order to determine the positive clones, use pointed tweezers to make several small holes (asymmetric distribution) in the filter membrane and culture medium to mark the position.
For more product details, please visit Aladdin Scientific website.